Breaking news
[Back to archive]
The latest results achieved by the project consortium.
Saccadic adapation modifies perceptual localization
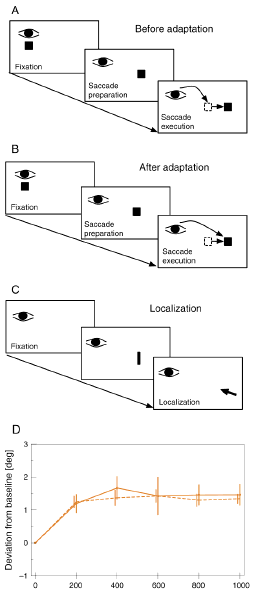
Fig 1: a) Artificial introduction of a post-saccadic visual error by stepping the target while the saccade is in flight. b) Adaptation of saccade behavior according to the post-saccadic error. c) Localization test after adaptation occurred. d) Saccade amplitude changes (dashed line) and localization changes (continuous line) over the course of adaptation.
Vision informs us about the location of objects around us. A large number of visual brain areas feature neurons with spatially selective receptive fields, which can in principle convey information about the location of visual stimuli. How does the brain deduce from these many possibilities where objects are located? From an ecological perspective, localization is plainly needed for behavior control. Eye movements are the most ubiquitous behavior, and if we want to know anything about an object somewhere in the scene we usually first look at it. Thus, the most pressing need of visual localization of any object is to guide eye movements to that object. It has been proposed that our perceptual experience of the world is really composed of the sensorimotor transformation laws that govern how we interact with the world (ORegan, J. K., & Noe, A. Behavioral Brain Science, 24, 9391031, 2001). This proposal predicts that our perception of the location of an object draws on the knowledge of the motor system about how to target this object with an eye movement.
We have used saccadic adaptation to modify the motor response to a target object. Normally, when a saccadic eye movement is initiated to an eccentric target the oculomotor system calculates the correct speed and duration of the movement in advance since, because of latencies in the visual system, visual feedback about the accuracy of the eye movement arrives in the brain only after the saccade is finished. Thus, saccades are 'ballistic' and often not very accurate. If a saccade happens to be inaccurate the error between the final gaze direction and the target can only be registered after the saccade. This post-saccadic visual error is used by the oculomotor system to check saccadic accuracy and, if needed, re-calibrate motor control for subsequent saccades to be more accurate. In the experimental paradigm of saccadic adaptation, a post-saccadic visual error is introduced artificially by stepping the target while the saccade is in flight (Figure 1A). Because vision is suppressed during saccades the step itself is not seen by the subject, but the oculomotor system registers the post-saccadic error and adapts saccade behavior accordingly: over the course of several such trials, the amplitude of the saccade to the target eventually matches the stepped location of the target, rather than the initial location (Figure 1B). The saccadic adaptation procedure, therefore, introduces a dissociation between the physical location of the target as registered by the retina and the motor program that is executed to move gaze onto that target. This dissociation allows us to investigate whether the perceptual localization of a stimulus at the target location follows the physical location of the stimulus or the motor program set-up by the saccadic system.
To decide this question we combined a series of adaptation trials with an interspersed localization task. The subject sat in front of a computer monitor which displayed a fixation target. The subject had to fixate the stimulus and initiate a saccade as soon it moved to the right. To induce saccadic adaption, the target stimulus was stepped a further 3 deg to the right when the eye tracker detected the onset of the saccade. This procedure was repeated for 1000 trials to achieve stable adaptation and allow the investigation of localization effects over the course of the modification of the saccade parameters.
After every 200 adaptation trials localization was tested in a block of 20 trials. Subjects were instructed to direct gaze at the center of the screen and avoid any eye movement during the entire block of localization trials (Figure 1C). A small localization probe was presented for 30 ms 13 deg to the right of the subjects current gaze position. The subject indicated the apparent location of the probe with the mouse pointer.
Figure 1D shows saccade amplitude changes (dashed line) and localization changes (continuous line) over the course of adaptation, starting from a pre-adapation baseline. Adaptation reached a maximal amplitude increase of 1.4 deg, or 46 % of the 3 deg target step. The localization also changed over the course of adaptation and showed a very similar development as the adaptation. After 1000 trials of adaptation localization probes were mislocalized 1.3 deg in the direction of adaptation. These results clearly show that modifications of saccadic amplitude by saccadic adaptation can induce associated changes in visual localization, consistent with the view that oculomotor knowledge is used in the perceptual representation of space.
Zimmermann E., Lappe M.
Department Psychology
University of Münster, Germany
|
